Stefan Helmreich, Anthropology, Massachusetts Institute of Technology
In How Like a Leaf, Donna Haraway, in interview with Thyrza Nichols Goodeve, reflects on her early interest in marine biology, reminiscing about a summer course she attended in 1968 at the Marine Biological Laboratory in Woods Hole, Massachusetts.[i] She recalls her fascination with sea grapes, a colonial marine organism able continually to regenerate its constituent parts, called zooids. Many years after her encounter with this totipotent tunicate, Haraway again meditated upon the generative capacities of marine creatures, this time in her foreword to Women Writing Culture, in which she turned to coral reefs and described “non-mammalian replicative doings among marine invertebrates: egg-release into open waters, followed by larval feeding, and finally the settling and metamorphosis into the adult forms of myriad species.” She likened the process of crafting conversation on common reading and writing to the process of reef building: “the written, collected, and published book of interviews becomes the finished scaffolding, the coraline reef, on which the next generations of spineless, non-bilaterally symmetrical entities will settle, eat each other and passers-by, and proliferate their drifting, always hungry, and seedy brood.”[ii]
In this essay, I fasten my attention on this figure of the reef. Haraway’s analyses have often been attached to figures — creatures of fact and fiction that symbolize and embody social and scientific tensions, trends, and transformations. Thus her cyborg, vampire, modest witness, FemaleMan, and OncoMouse™ are figures that gather up the ways that information technology, flows of blood, transfers of genes, and images from science fiction have rewired, reformatted, and rerouted many of our apprehensions and experiences of our bodies and selves. Drifting into this densely populated zone, I nominate the coral reef as a figure worthy of joining this Harawavian menagerie. Coral reefs, I propose, can attune their human visitors and inquisitors to empirical and epistemological questions of scale and context — where context, drawing upon a once-upon-a-time literal but now more figural meaning, refers to a “weaving together” (OED). Which earthly and oceanic entities and agents might be woven together through reefy bones and bodies is, of course, a polysemous, shifting question. Indeed, part of what I am riveted by when it comes to coral reefs is how densely they come prefigured through the historically layered descriptions of biologists, fisherpeople, ecologists, and, occasionally, members of my own professional guild, anthropology. Offering a reading of reefs, then, must simultaneously perform a kind of archaeology of the proliferative scaffoldings upon which reefs have already been written.
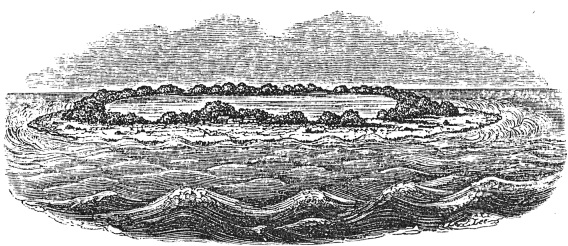
FIGURE 2

In How Like a Leaf, Donna Haraway, in interview with Thyrza Nichols Goodeve, reflects on her early interest in marine biology, reminiscing about a summer course she attended in 1968 at the Marine Biological Laboratory in Woods Hole, Massachusetts.[i] She recalls her fascination with sea grapes, a colonial marine organism able continually to regenerate its constituent parts, called zooids. Many years after her encounter with this totipotent tunicate, Haraway again meditated upon the generative capacities of marine creatures, this time in her foreword to Women Writing Culture, in which she turned to coral reefs and described “non-mammalian replicative doings among marine invertebrates: egg-release into open waters, followed by larval feeding, and finally the settling and metamorphosis into the adult forms of myriad species.” She likened the process of crafting conversation on common reading and writing to the process of reef building: “the written, collected, and published book of interviews becomes the finished scaffolding, the coraline reef, on which the next generations of spineless, non-bilaterally symmetrical entities will settle, eat each other and passers-by, and proliferate their drifting, always hungry, and seedy brood.”[ii]
In this essay, I fasten my attention on this figure of the reef. Haraway’s analyses have often been attached to figures — creatures of fact and fiction that symbolize and embody social and scientific tensions, trends, and transformations. Thus her cyborg, vampire, modest witness, FemaleMan, and OncoMouse™ are figures that gather up the ways that information technology, flows of blood, transfers of genes, and images from science fiction have rewired, reformatted, and rerouted many of our apprehensions and experiences of our bodies and selves. Drifting into this densely populated zone, I nominate the coral reef as a figure worthy of joining this Harawavian menagerie. Coral reefs, I propose, can attune their human visitors and inquisitors to empirical and epistemological questions of scale and context — where context, drawing upon a once-upon-a-time literal but now more figural meaning, refers to a “weaving together” (OED). Which earthly and oceanic entities and agents might be woven together through reefy bones and bodies is, of course, a polysemous, shifting question. Indeed, part of what I am riveted by when it comes to coral reefs is how densely they come prefigured through the historically layered descriptions of biologists, fisherpeople, ecologists, and, occasionally, members of my own professional guild, anthropology. Offering a reading of reefs, then, must simultaneously perform a kind of archaeology of the proliferative scaffoldings upon which reefs have already been written.
Haraway never conjures her figures whole; rather, they are always borrowings — from, for example, the iconography of the scientific revolution, the culture of U.S. militarism, the tides of feminist politics, and the fables of Christian salvation history. Her figures are entities she has repurposed for her own ends. The coral reefs I grapple with here are no different. They too come with durable, multiple, and porous inheritances. Emphasizing their surprising capacities to connect scales and contexts must attend to these historical holdfasts. This essay divides into three parts, which more or less chronologically follow figurations of coral from their emergence as nineteenth-century architectures of curiosity, to their fashioning as twentieth-century polymorphs inviting immersive and fleshy encounter, to their twenty-first-century rewriting as nodes in global genetic networks that might be read in light of biomedical economies and climatological emergency.
Emergence
Coral began its career in the scientific imagination as a boundary organism, an assemblage of flesh and stone that generated speculation about the boundaries of the living and nonliving. Early natural history and biology thought these zoophytes to be odd compounds of animal, vegetable, and mineral, and their classification confounded such figures as Linneaus. But their liminality was precisely what captivated such later thinkers as Robert Grant, Charles Darwin’s teacher at Edinburgh University, who saw coral as a missing link between plants and animals and who inspired Darwin’s 1842 book, The Structure and Distribution of Coral Reefs. Darwin’s questions about coral in this book turned out to be largely geological; he wanted to know how reefs were formed, and he postulated that reefs emerged from the sea as their supporting substructures submerged (see figure 1).[iii] The comparisons that motivated his thinking were less organic than they were architectural. In 1839, for example, in his Voyage of the Beagle, he rhapsodized, “We feel surprise when travelers tell us of the vast dimensions of the Pyramids and other great ruins, but how utterly insignificant are the greatest of these when compared to these mountains of stone accumulated by the agency of various minute and tender animals!”[iv]
FIGURE 1
Whitsunday Island, a “lagoon-island” or atoll in the south Pacific. This sketch of a ring of coral left behind by island subsidence is from Darwin 1842.
Whitsunday Island, a “lagoon-island” or atoll in the south Pacific. This sketch of a ring of coral left behind by island subsidence is from Darwin 1842.
Just two years later, social-evolutionist-to-be Lewis Henry Morgan wrote an essay on reefs, inaugurating his career-long fascination with the way living things transformed their earthly surroundings. His later meditations on the creative character of nonhuman mammals in his 1868 The American Beaver and His Works and his interest in Iroquois notions of the land as an ancestral living presence stemmed from attempting to understand the creation of life through imagery in Genesis; according to the Biblical creation tale, “earth [was] the medium from which the living things [were] created.”[v] The example of earth Morgan fixed on first, though, in his 1841 “Essay on Geology,” was the coral island:
These islands … are formed by the labours of millions of little insects, whose industry and ingenuity almost exceed belief. —It is but lately that any attention has been directed to these animals in a scientific manner and many questions relating to the nature of the animalcules and to the manner in which these islands are elevated above the level of the sea, are not as yet fully answered.[vi]
For Morgan, coral stood as a symbol of life emerging from and returning to geology and the sea. Coral animalcules, bridging the past and the future, were animated by the practice of building the world and bodies they inhabited, an activity that linked them to human collectives, who, for the Morgan who wrote Ancient Society in 1872, could be classed by their scale of technological achievement, which then determined for him their stage of social evolution.
Figurations of reefs as examples of the production of the inorganic through organic enterprise inspired anthropologist Alfred Kroeber in his 1948 essay “The Nature of Culture,” in which he used coral to illustrate his concept of the “superorganic.” Kroeber, a student of Franz Boas (whose doctoral thesis in geography and physics was about the color of seawater, as seen from above), enlisted reefs to explain the culture concept:
Figurations of reefs as examples of the production of the inorganic through organic enterprise inspired anthropologist Alfred Kroeber in his 1948 essay “The Nature of Culture,” in which he used coral to illustrate his concept of the “superorganic.” Kroeber, a student of Franz Boas (whose doctoral thesis in geography and physics was about the color of seawater, as seen from above), enlisted reefs to explain the culture concept:
A simile that may further help the realization of what culture is and how it works is that of a coral reef. Such a reef may be miles long and inhabited by billions of tiny polyp animals. The firm, solid part of the reef consists of calcium carbonate produced by the secretions of these animals over thousands of years — a product at once cumulative and communal and therefore social. What is alive and organic in the reef is these innumerable little animals on its ocean-fronting surface. Without their ancestors, there would have been no reef. But the reef now exists independently of the living polyps, and would long continue to endure even if every polyp were killed by, say, a change in ocean temperature or salinity. … While a coral reef is the accumulated precipitate of dead polyps, it is also a phenomenon affording to millions of living polyps a base and a foothold, and a place to thrive.[vii]
Kroeber concluded: “Each of us undoubtedly contributes something to the slowly but ever changing culture in which we live, as each coral contributes his gram or two of lime to the Great Barrier Reef.” What is remarkable about this description — aside from its sexing of polyps as males (a strict, if problematic, gametic reductionism would alert us to the fact that many coral host both eggs and sperm) — is the way Kroeber described a social constructivism that was enabled by biogenesis, even as the biotic part of the equation became finally less intriguing to him than the “superorganic.” As in Darwin’s and Morgan’s coral conceptions, what is compelling for Kroeber are not so much the organisms themselves, but the results of their social labor; that is, what emerges from their collective agency, not how the agency is itself constituted. These writers are primarily concerned with what Haraway, above, calls “the finished scaffolding” of cultural production, and are less interested in how “the next generations … will settle, eat each other and passers-by, and proliferate their drifting, always hungry, and seedy brood.”[viii]
Coral scaffolding was of interest to another kind of actor at mid-twentieth century: the United States military, which as part of its nuclear weapons testing projects, drilled deep into limestone reefs around Enewetak Atoll in the Marshall Islands in order to test the site’s geological stability for detonating the world’s first hydrogen bomb. The drilling that set the stage for the explosion had the unanticipated dividend of providing geological evidence for Darwin’s century-old theory of coral formation. More consequential for biology — particularly as lived experience — however, were the devastating effects on the South Pacific of what Ward Churchill and Winona LaDuke have called “radioactive colonialism.”[ix] Nuclear fallout and its egregious (mis)management by the United States radically transformed the social and ecological realities in this huge area of water, islands, and people, leaving a legacy of disease and environmental racism that persists to this day. Anthropologist Joe Masco has described how nuclear testing changes the way “nature” is imagined and inhabited by those haunted by the half-lives of radioactive elements. What he terms “mutant ecologies” — “enriched,” for example, with bomb carbon — have been woven into the microstructure of local biogenesis. “[T]he Bikini Island ecosystem” (where the US began atomic testing in 1946), for example, “continues to negotiate the eight-day half life of iodine-131, the 28.5-year half-life of strontium-90, and the 24,500-year half-life of plutonium.”[x] Such mortal negotiations show up as cancer in humans and radioactivity in coral reef fish.
One year after Enewetak, in 1953, Harper and Brothers published in English the first book by Jacques Cousteau, The Silent World, which included some of the first photos of fleshy life on the reef. SCUBA diving, along with the ecological aesthetic of writers such as Rachel Carson, would in the next decades transform the idioms within which oceanographers and others would figure reefs. In her 2010 ethnographic meditation on laboratory research into coral generation at the Long Marine Laboratory in Santa Cruz, California, Eva Hayward writes of the “fingeryeyes” that now materialize in the hydro-optics of marine research on cup coral.[xi] In the wake of Cousteau, the fingery tendrils of coral polyps would beckon scientists and their publics to new promises of what we can call, borrowing from anthropology, immersive participant-observation. Coral would become interesting not just as architecture, but as animate matter. Reefs would come to new life.
Coral scaffolding was of interest to another kind of actor at mid-twentieth century: the United States military, which as part of its nuclear weapons testing projects, drilled deep into limestone reefs around Enewetak Atoll in the Marshall Islands in order to test the site’s geological stability for detonating the world’s first hydrogen bomb. The drilling that set the stage for the explosion had the unanticipated dividend of providing geological evidence for Darwin’s century-old theory of coral formation. More consequential for biology — particularly as lived experience — however, were the devastating effects on the South Pacific of what Ward Churchill and Winona LaDuke have called “radioactive colonialism.”[ix] Nuclear fallout and its egregious (mis)management by the United States radically transformed the social and ecological realities in this huge area of water, islands, and people, leaving a legacy of disease and environmental racism that persists to this day. Anthropologist Joe Masco has described how nuclear testing changes the way “nature” is imagined and inhabited by those haunted by the half-lives of radioactive elements. What he terms “mutant ecologies” — “enriched,” for example, with bomb carbon — have been woven into the microstructure of local biogenesis. “[T]he Bikini Island ecosystem” (where the US began atomic testing in 1946), for example, “continues to negotiate the eight-day half life of iodine-131, the 28.5-year half-life of strontium-90, and the 24,500-year half-life of plutonium.”[x] Such mortal negotiations show up as cancer in humans and radioactivity in coral reef fish.
One year after Enewetak, in 1953, Harper and Brothers published in English the first book by Jacques Cousteau, The Silent World, which included some of the first photos of fleshy life on the reef. SCUBA diving, along with the ecological aesthetic of writers such as Rachel Carson, would in the next decades transform the idioms within which oceanographers and others would figure reefs. In her 2010 ethnographic meditation on laboratory research into coral generation at the Long Marine Laboratory in Santa Cruz, California, Eva Hayward writes of the “fingeryeyes” that now materialize in the hydro-optics of marine research on cup coral.[xi] In the wake of Cousteau, the fingery tendrils of coral polyps would beckon scientists and their publics to new promises of what we can call, borrowing from anthropology, immersive participant-observation. Coral would become interesting not just as architecture, but as animate matter. Reefs would come to new life.
Immersion
In the last half of the twentieth century, scientific figurations of coral shifted focus from bones to flesh — to pulpy polyp and algal bodies, brooding and spawning. To be sure, much of this interest might be chalked up to transformations in methods of study. Where Darwin and others primarily encountered dead coral fragments, and imagined these sculptural, sepulchral forms almost as archaeological artifacts, twentieth-century naturalists sought to submerge their bodies and eyes in the midst of coral communities (the closest Darwin came was a sojourn at low tide to visit outer corals in Tahiti with the aid of a “leaping pole.”[xii]). Architectural language still surfaces in descriptions of reef structures from time to time, albeit in a different register. Journalist Osha Davidson, in his loving 1998 account of coral reefs around the world, The Enchanted Braid, writes that seeing a reef underwater is “like being dropped into the center of a huge city on an alien planet.”[xiii] Coral cities are now alive, not archeological. And instead of being simple analogs to human cultural productions, they have become a more lively other (figure 2). We might say that they have become subjects for an appreciative relativism rather than for the derivation of universalistic lessons of the sort gleaned by Kroeber.

FIGURE 2
An immersive encounter with the fleshy coral other. “Penny Bailey observes coral spawning in the Florida Keys National Marine Sanctuary off Key Largo,” Florida Keys eNewsletter, http://www.fla-keys.com/newsletter/200508/
It is not difficult to argue, dipping into the anthropological inkwell once again, that corals have been figured as critters offering a cultural critique of received wisdom about the nature of such things as embodiment and sex. To begin with bodies: coral are classed as animals that live in “colonies” made of thousands of small sedentary creatures called coral polyps. A polyp is “a tiny ring of gelatinous tentacles fluttering above an equally small, internally rippled sac. Hard corals also have a skeleton, or corallite, at their base, into which the polyp retreats during the day.”[xiv] These polyps are symbiotic with a microscopic form of algae called zooxanthellae, which live inside the polyps and provide them with nutrients derived from photosynthesis.[xv] This relationship with zooxanthellae also helps to “speed up the process by which corals build their stony skeleton, putting down layer after layer of calcium carbonate, a form of limestone. It is this process of calcification that physically constructs coral reef, the literal bedrock of the coral reef.”[xvi] This mutualistic relation between an assemblage of animal and plant is a compound, what Haraway calls “a composite of individual organisms, an enclosure of zoons, a company of critters infolded into a one.”[xvii] On this view, coral can be a model for distributed subjectivities and agencies, for what anthropologist Marilyn Strathern, following McKim Marriot, has called “dividuality,” the partibility of persons or subjects.[xvii]
On sex and reproduction: Until the late 1970s, most marine biologists believed that coral was viviparous; that is, that polyps plopped up new polyps through an asexual process called budding. This model was called into question when biologists at James Cook University in Townsville, Australia in 1979 discovered that many coral “broadcast” sperm and eggs into the ambient waters around their reefs, producing slicks of gametes that provide yummy food for fish, but also allow for the creation of planktonic coral larvae (figure 3). In many settings, such spawning happens only once a year, on a full moon. When it was established that most coral spawned, and that larvae could survive for days, it became evident that widely distributed reef ecologies were hyperlinked to one another. Coral ecologies turned out to be translocally constituted — connected through ocean currents, which Davidson calls “watery highways for coral larvae.”[xix] The spawn of polyps in one locale are relations of those in another; more, in the process of recombination that unfolds during spawning, new sorts of corals can come into being through hybridization. Sexual relations among coral are distributed beyond discrete bodies and result not so much in populations of individuals, but in networks of dividuals. These webs of relation continually recontextualize one another.
On sex and reproduction: Until the late 1970s, most marine biologists believed that coral was viviparous; that is, that polyps plopped up new polyps through an asexual process called budding. This model was called into question when biologists at James Cook University in Townsville, Australia in 1979 discovered that many coral “broadcast” sperm and eggs into the ambient waters around their reefs, producing slicks of gametes that provide yummy food for fish, but also allow for the creation of planktonic coral larvae (figure 3). In many settings, such spawning happens only once a year, on a full moon. When it was established that most coral spawned, and that larvae could survive for days, it became evident that widely distributed reef ecologies were hyperlinked to one another. Coral ecologies turned out to be translocally constituted — connected through ocean currents, which Davidson calls “watery highways for coral larvae.”[xix] The spawn of polyps in one locale are relations of those in another; more, in the process of recombination that unfolds during spawning, new sorts of corals can come into being through hybridization. Sexual relations among coral are distributed beyond discrete bodies and result not so much in populations of individuals, but in networks of dividuals. These webs of relation continually recontextualize one another.
FIGURE 3
Coral egg and sperm bundles released in spawning. From Gilmour 1999. See “Photograph of the week: 07 February 2001,” FNAS School of Animal Biology 2001-Feb-07, http://www.animals.uwa.edu.au/latest/2001/2001feb07
Perhaps the biggest stars of the reef when it comes to offering examples of sexual variety, however, have been the sequentially hermaphroditic fish that live in the embrace of many coral reefs. These are fish that can change “sex” (the production of distinctive hormones, kinds of gametes, and scaly coloring being the markers here) in response to social cues like the balance of hormones in their ambient crowd of compatriots. Such piscines — by and large transitioning FTM — have been favorite creatures for cultural critiques of the stability of natural sex identity, reminiscent of the kinds of biological differences offered by biologist Anne Fausto-Sterling in her argument that, when it comes to apportioning sexual identity based on genes, chromosomes, genitals, and hormones, we might reasonably splice the world into at least “five sexes” rather than the traditional two.[xx] Evolutionary ecologist Joan Roughgarden in Evolution’s Rainbow, reaching into reef ecologies, discusses coral bass that switch within one mating episode between making sperm and making eggs. While she draws instructively upon her own experience as an MTF human, much of Roughgarden’s argumentation works sociobiologically, naturalizing varieties of human behavior either by anchoring them in long-ago adaptations or by comparing them to purportedly analogous activities in other animals. Her definition of sex — based on the size of gametes an organism produces — is curiously reductionist for someone ostensibly so attuned to the variety of scales and contexts within which something so unruly and vague as “sex”(as practice, as identification, as trajectory) materializes. Elizabeth Wilson, revisiting Darwin’s work on barnacles and his puzzlement over their polymorphous sexual arrangements, tries to be more cautious in drawing lessons about sexuality from marine organisms. In her “Biologically Inspired Feminism,” she writes that,
to characterize Darwin’s barnacles as queer is too glib — if by this characterization we mean that the barnacle simply mimics those human, cultural, and social forms now routinely marked queer (the transgender barnacle! the polyandrous barnacle!). This characterization has more punch if it is used, contrariwise, to render those familiar human, cultural, and social forms more curious as a result of their affiliation with barnacle organization.[xxi]
In other words, treating a creature like coral as a figure rather than a sedimented site of literal truths about nonhuman nature is a better bet for generating a socially useful discourse on biology. Eva Hayward’s 2010 essay on Balanophyllia elegans, cup corals, offers a state-of-the-art-and-science example of how to “disarticulate sex, sexuality, and reproduction” and then, perversely, to rearticulate them with a diffractive attention to “perverting meaning, inverting power, gendered perversions, coralline inversions.” “Perhaps both species of ‘inverts,’” writes Hayward, “the kind without backbones and the sort who transpose gender roles, interrupt heteronormativity, although not for the same reasons, but because of a shared activity of making bodies pliable, mobile, transposable.”[xxii] Coral is good to queer with.
Belleville, Bill
Kroeber, Alfred L.
Emergency
If architectural metaphors had coral as a primitive premonition of the more filigreed cultural construction projects of humans and if attention to coral sex and reproduction has sometimes served as a reminder that sex/gender does not describe a natural hierarchy or binary, how is the world of the coral reef predominantly figured now, at the opening of the twenty-first century? In the wake of the 1992 Rio summit, reefs became a symbol for marine biodiversity. Indeed, it is now easy to find descriptions of coral reefs as “rainforests of the sea.” And like rainforests, they are under threat.
Recent worldwide degradation of coral reefs has been linked to the effects of too much carbon dioxide in the atmosphere — that is, to global warming, which can make sea levels rise faster than the pace of coral growth. When this happens, the symbiotic algae in coral polyps lose the ability to photosynthesize and may be ejected from their hosts in a process called “bleaching,” so named because it leaves polyps translucent, allowing white coral bone to show through. Corals also suffer when sunlight is obstructed by the rapid growth of surface algae, which often happens in water suddenly loaded with nutrients from sewage flow or fertilizer run-off (think coastal golf courses). Because of the calibration of these deleterious changes to large-scale processes, many ecologists now consider coral to offer a measure of planetary health. Because the changes registered indicate declining oceanic health, and are often triggered by anthropogenic causes, scientists often pitch coral as delivering a message from the living planet: “Coral reefs may be warning us to pay closer attention, just as they can signal the pressures that modern populations are placing on tropical resources.”[xxiii] In their barometric readings, reefs sound a warning signal from Gaia, chastising humans for self-indulgent, shortsighted activities.
Environmentally conscious scientists often translate this scold into one particularly aimed at so-called developing, hyper-urbanized countries such as the Philippines or Indonesia, yet to tune into Gaia’s alert.[xxiv] As participants at an international conference on coral reefs held in Bali in October 2000 argued, however, reef degradation must be seen through the lens of present day political economy.[xxv] The depredation of, say, the Java Sea — flooded with runoff from logging and agriculture; mined for coral construction materials; blast fished for food; and scoured for tropical fish for international collectors — is linked to the dynamics and demands of intercalated and unequal local and global markets.[xxvi] Reefs are not just climatic barometers but also serve as indicators of North-South inequality. The contexts they conjoin are legibly political, connecting local biogenesis to international patterns of fossil fuel usage, tourism, and food production. In worries about climate change, the flow and connection that captured the attention of coral biologists in the 1970s and 80s has been eclipsed by a sense that coral reefs are under threat because of their immobility; they drown or bleach because they cannot move. Flipping prefixes in a phrase from Bruno Latour, they are mutable immobiles.[xxvii]
Any figuration, of course, is prone to gestalt shifts in what will count as figure and ground, and coral immobility has also been construed as a virtue. Coral’s defense against predators comes not from the evasive mobility so characteristic of many fish, but from the manufacture of toxins that can be released when polyps come under threat. This has made coral an intriguing creature for natural products chemists interested in drug leads. The Coral Reef Research Foundation (CRRF), founded in 1991 by Patrick Colin and Lori Bell and supported in part by the United States National Cancer Institute (NCI), has employed scientists to collect marine invertebrates as potential sources for new anticancer drugs (see http://www.coralreefresearchfoundation.org/index.html). CRRF, a non-profit organization, is incorporated in California but also in the Indo-Pacific Island nation of Palau, which became independent from the U.S. in 1994. CRRF scientists freeze and fly material samples to the headquarters of the NCI in Maryland, where they are screened for bioactivity against cancer cells and HIV. Recognizing Palauan sovereignty over of these resources, the NCI has in place a bioprospecting contract with Palau. If the NCI wants compounds sourced in Palau to be developed into drugs, the contract specifies that Palau must be compensated by any pharmaceutical partners to whom the U.S. patent is licensed. On paper, this contract seeks to ameliorate North-South inequalities by putting a price on coral biodiversity. Whether this agreement secures a flow of benefits is not clear; a study of the sort undertaken by Cori Hayden in When Nature Goes Public, a 2003 ethnographic account of a bioprospecting contract between the National Autonomous University of Mexico and the University of Arizona (partnered with Wyeth-Ayerst and American Cyanamid) would be instructive.
These most recent figurations of coral — as a monitor of planetary health and a potential resource for tropical economic health and biomedical human health — are threaded through — what else? — genomics.[xxviii] It should come as no surprise that many coral biologists have lately called for a “Coral Genome Project.” “Coral-list,” a digital scaffolding of online conversation scrabbled together, in English, mostly by coral researchers, is one place to look for particulars.
Almost as soon as the subject of a sequenced coral genome was raised on Coral-list, talk turned to the question of which genus of reef-building coral might offer the most useful model system. Scientists engaged in practices of figuration as they discussed which coral DNA to sequence. The first creature floated as a candidate was the reef-building lobe coral, Porites lobata — which, its advocates maintained, showed “rising importance as a ‘laboratory rat’ in coral exotoxicology, coral cell biology, coral immunity and coral neurophysiology,” and might be used as “a model for molecular genetics, cell biology, biochemistry, lipid chemistry, sterol/polyphenol chemistry, environmental/physiological monitoring, ecotoxicology, stress physiology, coral immunity, coral oncology, coral endocrinology and coral neurophysiology.”[xxix] Its wide distribution in the Indian and Pacific Oceans, the Red Sea, and Persian Gulf was also offered in its favor, though its durability, flexibility, and amenability to molecular and biochemical techniques in the laboratory setting were selling points as well. Porites was lab-friendly because, as a summary of the discussion had it, “Nucleic acid isolation and in situ hybridization and RNAi would work better the ‘meatier’ the coral.” Even in this new realm of code, flesh matters (see figure 4). Another consideration in choosing a species would be how well it allowed for study of the interaction between polyps and their mutualist endosymbionts, the zooxanthellae. This could be instructive for understanding coral bleaching. A 2004 New York Times article suggested that some robust strains of “heat-tolerant algae may move in to replace strains lost in bleaching events.”[xxx]



FIGURE 6
Notes
Recent worldwide degradation of coral reefs has been linked to the effects of too much carbon dioxide in the atmosphere — that is, to global warming, which can make sea levels rise faster than the pace of coral growth. When this happens, the symbiotic algae in coral polyps lose the ability to photosynthesize and may be ejected from their hosts in a process called “bleaching,” so named because it leaves polyps translucent, allowing white coral bone to show through. Corals also suffer when sunlight is obstructed by the rapid growth of surface algae, which often happens in water suddenly loaded with nutrients from sewage flow or fertilizer run-off (think coastal golf courses). Because of the calibration of these deleterious changes to large-scale processes, many ecologists now consider coral to offer a measure of planetary health. Because the changes registered indicate declining oceanic health, and are often triggered by anthropogenic causes, scientists often pitch coral as delivering a message from the living planet: “Coral reefs may be warning us to pay closer attention, just as they can signal the pressures that modern populations are placing on tropical resources.”[xxiii] In their barometric readings, reefs sound a warning signal from Gaia, chastising humans for self-indulgent, shortsighted activities.
Environmentally conscious scientists often translate this scold into one particularly aimed at so-called developing, hyper-urbanized countries such as the Philippines or Indonesia, yet to tune into Gaia’s alert.[xxiv] As participants at an international conference on coral reefs held in Bali in October 2000 argued, however, reef degradation must be seen through the lens of present day political economy.[xxv] The depredation of, say, the Java Sea — flooded with runoff from logging and agriculture; mined for coral construction materials; blast fished for food; and scoured for tropical fish for international collectors — is linked to the dynamics and demands of intercalated and unequal local and global markets.[xxvi] Reefs are not just climatic barometers but also serve as indicators of North-South inequality. The contexts they conjoin are legibly political, connecting local biogenesis to international patterns of fossil fuel usage, tourism, and food production. In worries about climate change, the flow and connection that captured the attention of coral biologists in the 1970s and 80s has been eclipsed by a sense that coral reefs are under threat because of their immobility; they drown or bleach because they cannot move. Flipping prefixes in a phrase from Bruno Latour, they are mutable immobiles.[xxvii]
Any figuration, of course, is prone to gestalt shifts in what will count as figure and ground, and coral immobility has also been construed as a virtue. Coral’s defense against predators comes not from the evasive mobility so characteristic of many fish, but from the manufacture of toxins that can be released when polyps come under threat. This has made coral an intriguing creature for natural products chemists interested in drug leads. The Coral Reef Research Foundation (CRRF), founded in 1991 by Patrick Colin and Lori Bell and supported in part by the United States National Cancer Institute (NCI), has employed scientists to collect marine invertebrates as potential sources for new anticancer drugs (see http://www.coralreefresearchfoundation.org/index.html). CRRF, a non-profit organization, is incorporated in California but also in the Indo-Pacific Island nation of Palau, which became independent from the U.S. in 1994. CRRF scientists freeze and fly material samples to the headquarters of the NCI in Maryland, where they are screened for bioactivity against cancer cells and HIV. Recognizing Palauan sovereignty over of these resources, the NCI has in place a bioprospecting contract with Palau. If the NCI wants compounds sourced in Palau to be developed into drugs, the contract specifies that Palau must be compensated by any pharmaceutical partners to whom the U.S. patent is licensed. On paper, this contract seeks to ameliorate North-South inequalities by putting a price on coral biodiversity. Whether this agreement secures a flow of benefits is not clear; a study of the sort undertaken by Cori Hayden in When Nature Goes Public, a 2003 ethnographic account of a bioprospecting contract between the National Autonomous University of Mexico and the University of Arizona (partnered with Wyeth-Ayerst and American Cyanamid) would be instructive.
These most recent figurations of coral — as a monitor of planetary health and a potential resource for tropical economic health and biomedical human health — are threaded through — what else? — genomics.[xxviii] It should come as no surprise that many coral biologists have lately called for a “Coral Genome Project.” “Coral-list,” a digital scaffolding of online conversation scrabbled together, in English, mostly by coral researchers, is one place to look for particulars.
Almost as soon as the subject of a sequenced coral genome was raised on Coral-list, talk turned to the question of which genus of reef-building coral might offer the most useful model system. Scientists engaged in practices of figuration as they discussed which coral DNA to sequence. The first creature floated as a candidate was the reef-building lobe coral, Porites lobata — which, its advocates maintained, showed “rising importance as a ‘laboratory rat’ in coral exotoxicology, coral cell biology, coral immunity and coral neurophysiology,” and might be used as “a model for molecular genetics, cell biology, biochemistry, lipid chemistry, sterol/polyphenol chemistry, environmental/physiological monitoring, ecotoxicology, stress physiology, coral immunity, coral oncology, coral endocrinology and coral neurophysiology.”[xxix] Its wide distribution in the Indian and Pacific Oceans, the Red Sea, and Persian Gulf was also offered in its favor, though its durability, flexibility, and amenability to molecular and biochemical techniques in the laboratory setting were selling points as well. Porites was lab-friendly because, as a summary of the discussion had it, “Nucleic acid isolation and in situ hybridization and RNAi would work better the ‘meatier’ the coral.” Even in this new realm of code, flesh matters (see figure 4). Another consideration in choosing a species would be how well it allowed for study of the interaction between polyps and their mutualist endosymbionts, the zooxanthellae. This could be instructive for understanding coral bleaching. A 2004 New York Times article suggested that some robust strains of “heat-tolerant algae may move in to replace strains lost in bleaching events.”[xxx]
FIGURE 4
Sampling DNA from a Porites coral. “Coral Reefs May Have Fighting Chance of Surviving Climate Change,” Wildlife Conservation Society Reports: A Media Resource, http://www.wcs.org/wcspubs/wcsreports
Some advocates of Porites said that the huge coral heads this genus forms are good sources for records of long-term climate change — though there were also those who “didn’t agree that a coral that is primarily useful for fossil/paleoclimate studies would be one of the best choices for a genomics project,” arguing rather that “the coral community should figure out what would be the coral equivalent of a … fruit fly. A species should be selected that is most amenable to laboratory manipulations.” A feature of Porites that makes them potentially attractive lab creatures is that many do not spawn, but rather brood, releasing larvae throughout the year, rather than slicks of gametes only on particular nights.
Against Porites, others championed coral in the genus Acropora, the most widely distributed (and studied) genus in the world, which would in the bargain allow for a more thorough representation of Caribbean reefs (see figure 5). One participant added an intriguing pitch, arguing that Acropora arabia from Kuwait would be a fruitful subject for study because it seems to be tolerant of high temperature variations, salinities, turbidity, and — a regionally relevant geopolitical hazard — the presence of petroleum hydrocarbons swirling on the surface. The robustness of Acropora arabia, it was offered, might hold lessons for how “to help other fast-growing acroporids to maintain their distribution better.” On this view, coral must change with the times, whether it is in environments modulated by petroleum, global warming, or, in the South Pacific, radiation. Apropos of the Pacific thermonuclear tests in the 1950s, some biologists have begun to claim that the reefs around Bikini are recovering in “a testament to nature’s ability to heal itself.”[xxxi]
Against Porites, others championed coral in the genus Acropora, the most widely distributed (and studied) genus in the world, which would in the bargain allow for a more thorough representation of Caribbean reefs (see figure 5). One participant added an intriguing pitch, arguing that Acropora arabia from Kuwait would be a fruitful subject for study because it seems to be tolerant of high temperature variations, salinities, turbidity, and — a regionally relevant geopolitical hazard — the presence of petroleum hydrocarbons swirling on the surface. The robustness of Acropora arabia, it was offered, might hold lessons for how “to help other fast-growing acroporids to maintain their distribution better.” On this view, coral must change with the times, whether it is in environments modulated by petroleum, global warming, or, in the South Pacific, radiation. Apropos of the Pacific thermonuclear tests in the 1950s, some biologists have begun to claim that the reefs around Bikini are recovering in “a testament to nature’s ability to heal itself.”[xxxi]
FIGURE 5
Indicators of evolutionary relationships among Acropora. “Mean Base Compositions (%) of the Pax-C Intron and the mtDNA Intergenic Region (both repeat and nonrepeat regions) in Acropora Species.” From van Oppen, Madeleine J. H., Brenda J. Mcdonald, Bette Willis and David J. Miller (2001).
Questions of figuration were central in this Coral-list discussion. Environmental representativeness, lab tractability, and conservation concerns were all in the mix, channeled through gene talk. Biomedical interest was not absent from the conversation; indeed, the summary from which I have been quoting concluded that
Basic science questions can be greatly aided by knowing the sequence of the coral genome. For example, is this coral immunocompetent or endocrine modulated? We need to know the genes that contribute to these systems to explore their individual and combined behaviors…. Can our understanding of cancer in mammals be aided by our understanding and the future discoveries of how corals get cancer?
Some participants questioned the limits of the figuration work that coral genomics could do. One pointed out that “Corals are probably not a monophyletic group and the different families differ significantly in their physiology, ecology, and genetics.” But coral biodiversity can fit neatly into other genres of figuration. Recent work in coral genomics has shown coral and humans sharing genes bound up in the nervous system — genes intriguingly missing from the standard model organism of Drosophila — a result that could press for a rethinking of the common ancestor of coral and animals.[xxxii]
In the figurations of coral I have followed here, I discern a movement from opacity, to visibility, to readability. For Darwin, coral were glimpsed dimly, as bare bone, after death and, if living, through foamy water. For twentieth-century SCUBA diving scientists, coral were best encountered from an immersive, bodied, point of view. And for today’s environmentalists, biotechnologists, and would-be coral genomicists, coral are something to be read — for climate change, for potentially patentable genes, for representativeness. This itinerary of signifying substance — bones, flesh, genome — might be examined alongside the tripartite division that Haraway offered in her history of biological kinship categories in the United States, “Universal Donors in a Vampire Culture,” in which she suggested that craniometry, blood typing, and gene mapping have been successive techniques for figuring human (racialized) difference.[xxxiii] What difference does a figuration of coral make? What successor epistemologies can we imagine settling on and metamorphosing the textual idioms of genomics?
Building on Haraway’s comparison of reading to reef building, I want to suggest that the idea of reading coral represents an opportunity for reconstructing our understanding of the cultural and scientific politics of the sea — or, better, for reconfiguring such understandings, since coral must be read not simply or only as a sign, but as a figure.
In the figurations of coral I have followed here, I discern a movement from opacity, to visibility, to readability. For Darwin, coral were glimpsed dimly, as bare bone, after death and, if living, through foamy water. For twentieth-century SCUBA diving scientists, coral were best encountered from an immersive, bodied, point of view. And for today’s environmentalists, biotechnologists, and would-be coral genomicists, coral are something to be read — for climate change, for potentially patentable genes, for representativeness. This itinerary of signifying substance — bones, flesh, genome — might be examined alongside the tripartite division that Haraway offered in her history of biological kinship categories in the United States, “Universal Donors in a Vampire Culture,” in which she suggested that craniometry, blood typing, and gene mapping have been successive techniques for figuring human (racialized) difference.[xxxiii] What difference does a figuration of coral make? What successor epistemologies can we imagine settling on and metamorphosing the textual idioms of genomics?
Building on Haraway’s comparison of reading to reef building, I want to suggest that the idea of reading coral represents an opportunity for reconstructing our understanding of the cultural and scientific politics of the sea — or, better, for reconfiguring such understandings, since coral must be read not simply or only as a sign, but as a figure.
FIGURE 6
Margaret Wertheim leads a Hyperbolic Crochet Coral Reef workshop at NYU. Photo, Nicholas Roberts, New York Times, March 4, 2008.
Coral figures significantly for the Los Angeles-based Institute for Figuring (IFF), “an organization dedicated to the poetic and aesthetic dimensions of science, mathematics and the technical arts.”[xxxiv] Coordinating community-based and community-crossing handicraft projects, the IFF’s co-directors, Margaret and Christine Wertheim, seek to promote and facilitate hands-on apprehension of such scientific and natural forms as the Fibonacci series, the snowflake, and the sea slug through the modeling and making work of weaving, knitting, and origami — a fusion of calculation and fabrication that, according to a New York Times profile, threads together the Wertheims’ interests in “science, mathematics, art, feminism, handicrafts and social activism.”[xxxv] The activity for which the IFF has become best known is the Hyperbolic Crochet Coral Reef Project, a networked crocheting enterprise inaugurated in 2005 that means to create a material homage to the Great Barrier Reef through concatenating “loopy ‘kelps,’ fringed ‘anemones,’ and curlicued ‘corals’” fashioned through the craft of hyperbolic crochet, a technique invented by Cornell mathematician Daina Taimina in 1997 to model hyperbolic geometry in three-dimensions. As the Wertheims explain the enterprise, “The Institute For Figuring is crocheting a coral reef: a woolly celebration of the intersection of higher geometry and feminine handicraft, and a testimony to the disappearing wonders of the marine world.” The reef is already some 3,000 square feet in size, and has been exhibited and extended at events in Pittsburgh, Chicago, and New York. It includes such features as “The Ladies’ Silurian Atoll, a ring-shaped installation with close to 1000 individual crochet forms made by dozens of contributors from around the world.”[xxxvi] The New York Times calls the reef an “environmental version of the AIDS quilt,”[xxxvii] meant to draw attention to the worldwide destruction of reef systems, a concern dear to the Queensland-raised Wertheim sisters. Science studies scholar Sophia Roosth, who studied the making of this reef ethnographically, argues that new ways of figuring biology may be in the making here, ways that render “tangible the traces of gestural knowledge necessary to gather and sediment information about living forms.”[xxxviii] In parallel with the reading practices and politics of the Coral Genome Project, then, emerge the wooly writing activities of the Hyperbolic Crochet Coral Reef Project.
In her first book, Crystals, Fabrics, and Fields, Haraway examined how metaphor shapes scientific theory.[xxxix] Metaphor — from the Greek metafora, to “carry between” — transports conceptions from one realm to another. Figures, on the other polyp, direct a denser rhetorical traffic, spawning not only multiplicities of meanings, but also new material, corporeal structures and substances. Like coral: architected, embodied, experienced, read, written, rewritten, and woven into the fiber of our words and worlds.
In her first book, Crystals, Fabrics, and Fields, Haraway examined how metaphor shapes scientific theory.[xxxix] Metaphor — from the Greek metafora, to “carry between” — transports conceptions from one realm to another. Figures, on the other polyp, direct a denser rhetorical traffic, spawning not only multiplicities of meanings, but also new material, corporeal structures and substances. Like coral: architected, embodied, experienced, read, written, rewritten, and woven into the fiber of our words and worlds.
Acknowledgments: I thank Sharon Ghamari-Tabrizi for her invitation to think through coral using the figurations of Donna Haraway, whose research and writing has by now become so essential to the way I think. I have benefited, in reweavings, from the comments of Eva Hayward, Heather Paxson, Sophia Roosth, and Malcolm Shick. I am grateful to Katie King for making the Haraway Webfestschrift a real entity in our webby world.
Notes
[i] Haraway and Goodeve (2000). And see Haraway (1975).
[ii] Haraway (1995: xi-xii).
[iii] See Dobbs (2005) for a discussion of Darwin’s debate with Alexander Agassiz about the origin of reefs.
[iv] Quoted in Davidson (1998: 7).
[v] Feeley-Harnik (1999: 227).
[vi] Morgan, quoted in Feeley-Harnik (1999: 228).
[vii] Kroeber (1948: 255-256).
[viii] Haraway (1995: xi-xii).
[ix] Churchill and LaDuke (1992).
[x] Masco (2004: 525).
[xi] Hayward (2010).
[xii] Darwin (1842: 6).
[xiii] Davidson (1998: 55).
[xiv] Davidson (1998:14).
[xv] This relationship with zooxanthellae also helps to “speed up the process by which corals build their stony skeleton, putting down layer after layer of calcium carbonate, a form of limestone. It is this process of calcification that physically constructs coral reef, the literal bedrock of the coral reef” (Davidson 1998: 17).
[xvi] Davidson (1998: 17).
[xvii] Haraway (2004a: 3).
[xviii] See Strathern (1988).
[xix] Davidson (1998: 79).
[xx] Fausto-Sterling (1993).
[xxi] Wilson (2002: 284).
[xxii] Hayward (2010: 589).
[xxiii] Chadwick (1999: 37).
[xxiv] See Davidson (1998).
[xxv] See Moosa, et al. (2002).
[xxvi] Davidson (1998).
[xxvii] See Latour (1987) on immutable mobiles.
[xxviii] Though coral-as-bone still makes an appearance from time to time. The Bio-Eye® Orbital Implants website, which sells artificial eyes poses coral as a “natural choice” for ocular implants: “the goal of a more natural appearance was finally achieved with the help of a natural material: ocean coral. A remarkable similarity was noticed between the porous structure of certain coral species and that of human bone. … The eye muscles can be attached directly to this implant, allowing it to move within the orbit — just like the natural eye.”(“Bio-Eye® Orbital Implants,” http://www.ioi.com/patient/ha.html). “Nature” is crucial to popular rhetoric: “The stony skeletons of corals seem to be far more natural for bone and eye implants than artificial substances such as glass. The chemical composition of the coral, secreted from the minerals of the sea, is nearly identical to the complex mineral arrays created by the human body” (Belleville 1999:92). Here human kinship with the sea finds a link through the shared substance of hydroxyapatite, the complex calcium-phosphate salt found in human bones and coral skeletons. Future medicine promises to reverse the logic of Shakespeare’s famous line from The Tempest: “of his bones are coral made,” as coral becomes an organ donor for humans.
[xxix] All quotations are from a summary document produced by the Coral-list, “Summary of Professional Exchanges on Coral Genomics,” http://www.coris.noaa.gov/exchanges/coralgenome/sup_digest.html
[xxx] Dean (2004). On the other hand, another consideration “is the availability of zooxanthellae-free tissues. DNA from the zooxanthellae is often amplified (and subsequently sequenced) in addition to the coral DNA, unless the tissue is free from zooxanthellae or the primers are specific to cnidarians. The symbiotic dinoflagellates have a genome size which is estimated to be 100 times larger than the coral genome, further complicating the effort.”
[xxxi] Zorpette (1998: 24).
[xxxii] Kortschak, Samuel, Saint, and Miller (2003).
[xxxiii] Haraway (1994).
[xxxiv] The Institute for Figuring, http://www.theiff.org/
[xxxv] Cohen (2008).
[xxxvi] The Institute for Figuring, “About the Crochet Coral Reef,” http://crochetcoralreef.org/
[xxxvii] Cohen (2008).
[xxxviii] Roosth, personal communication. And see Roosth (2010).
[xxxix] Haraway (2004b).
References
Belleville, Bill
1999. Healing from the Sea. American Way, April 1:88-92.
Chadwick, Douglas H.
1999. Coral in Peril. National Geographic 195(1): 30-37.
Churchill, Ward and Winona LaDuke
1992. Native North America: The Political Economy of Radioactive Colonialism. In The State of Native America: Genocide, Colonization, and Resistance. M. Annette Jaimes, ed. Boston: South End Press, 241-266.
Cohen, Patricia
2008. Want to Save a Coral Reef? Bring along your Crochet Hook. New York Times, March 8, http://www.nytimes.com/2008/03/04/arts/design/04crochet.html?partner=rssnyt&emc=rss
Cousteau, Jacques, with Frédéric Dumas
1953. The Silent World. New York: Harper and Brothers.
Darwin, Charles
1842. The Structure and Distribution of Coral Reefs, reprint edition, 1984. Tucson: University of Arizona Press.
Davidson, Osha Gray
1998. The Enchanted Braid: Coming to Terms with Nature on the Coral Reef. New York: John Wiley and Sons, Inc.
Dean, Cornelia
2004. As the Seas Warm, Algae Help Some Coral Stand up to the Heat. New York Times, December 21, http://www.nytimes.com/2004/12/21/science/21cora.html
Dobbs, David
2005. Reef Madness: Charles Darwin, Alexander Agassiz, and the Meaning of Coral. New York: Pantheon Books.
Fausto-Sterling, Anne
1993. The Five Sexes: Why Male and Female Are Not Enough. The Sciences, March-April: 20-24.
Feeley-Harnik, Gillian
1999. “Communities of Blood”: The Natural History of Kinship in Nineteenth-Century America. Comparative Studies in Society and History 41(2): 215-262.
Gilmour, James P.
1999. An Experimental Investigation into the Effects of Suspended Sediment on Fertilisation, Larval Survival and Settlement in a Scleractinian Coral. Marine Biology 135: 451-462.
Haraway, Donna
1975. Marine Biological Laboratory of Woods Hole: An Ideology of Biological Expansion. American Academy of Arts and Sciences symposium on Knowledge in American Society, 1870-1920.
1994. Universal Donors in a Vampire Culture, Or It’s All in the Family: Biological Kinship Categories in the Twentieth-Century United States. In Uncommon Ground: Toward Reinventing Nature. William Cronon, ed., pp. 321-366. New York: Norton.
1995. Foreword to Women Writing Culture, Gary A. Olson and Elizabeth Hirsch, eds., pp. x-xii. Albany: State University of New York Press.
2000. How Like a Leaf, An Interview with Thyrza Nichols Goodeve. New York: Routledge.
2004a. Crittercam: Compounding Eyes in NatureCulture. Manuscript.
2004b. Crystals, Fabrics, and Fields: Metaphors that Shape Embryos. Foreword by Scott F. Gilbert. Reprint Version, Berkeley: North Atlantic Books (first edition, 1976).
Hayden, Cori
2003. When Nature Goes Public: The Making and Unmaking of Bioprospecting in Mexico. Princeton: Princeton University Press.
Hayward, Eva
2010. Fingeryeyes: Impressions of Cup Corals. Cultural Anthropology 25(4): 577-599.
Kortschak, R. Daniel, Gabrielle Samuel, Robert Saint, and David J. Miller.
2003. EST Analysis of the Cnidarian Acropora millepora Reveals Extensive Gene Loss and Rapid Sequence Divergence in the Model Invertebrates. Current Biology 13: 2190-2195.
Kroeber, Alfred L.
1948. Anthropology: Race, Language, Culture, Psychology, Prehistory, Revised edition. New York: Harcourt, Brace and Company.
Latour, Bruno
1987. Science in Action: How to Follow Scientists and Engineers through Society. Cambridge, MA: Harvard University Press.
Masco, Joe
2004. Mutant Ecologies: Radioactive Life in Post-Cold War New Mexico. Cultural Anthropology 19(4): 517-550.
Moosa, M.K. Kasim, S. Soeodihardjo, A. Nontji, A. Soegiarto, K. Romimohtarto, Sukarno and Suharsono, eds.
2002. Proceedings of the Ninth International Coral Reef Symposium, Bali, Indonesia, October 23-27, 2000. Ministry of Environment, Indonesian Institute of Sciences and the International Society for Reef Studies.
Roosth, Sophia
2010. Crafting Life: A Sensory Ethnography of Fabricated Biologies. Dissertation, Doctoral Program in History | Anthropology | Science, Technology, and Society, Massachusetts Institute of Technology.
Roughgarden, Joan
2004. Evolution's Rainbow: Diversity, Gender, and Sexuality in Nature and People. Berkeley: University of California Press.
Strathern, Marilyn
1988. The Gender of the Gift: Problems with Women and Problems with Society in Melanesia. Berkeley: University of California Press.
van Oppen, Madeleine J. H., Brenda J. McDonald, Bette Willis and David J. Miller.
2001. The Evolutionary History of the Coral Genus Acropora (Scleractinia, Cnidaria) Based on a Mitochondrial and a Nuclear Marker: Reticulation, Incomplete Lineage Sorting, or Morphological Convergence? Molecular Biology and Evolution 18: 1315-1329.
Wilson, Elizabeth
2002. Biologically Inspired Feminism. Australian Feminist Studies 17(39): 283-285.
Zorpette, Glenn
1998. Bikini’s Nuclear Ghosts. Scientific American Presents 9(3): 24-27.
Author Bio
Stefan Helmreich is Associate Professor of Anthropology at MIT. His research examines the work of biologists concerned with the conceptual and material limits of the category of “life.” Silicon Second Nature: Culturing Artificial Life in a Digital World (University of California Press, 1998) is an ethnography of theoretical biologists who use simulation techniques to model real and possible life forms. Alien Ocean: Anthropological Voyages in Microbial Seas (University of California Press, 2009) examines the lives of extremophilic marine microbes and the scientists who love them.
Web Design:
The website background illustration, "Niphates callista (Tube sponge) with sea cucumbers (Synaptula) and cup corals," photographed by Nick Hobgood and shared under a creative commons license, is from Wikipedia Commons: http://commons.wikimedia.org/wiki/File:Niphates_callista_%28Tube_sponge%29_with_sea_cucumbers_and_cup_corals.jpg{kind=link}